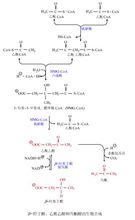
(1)酮体易运输:长链脂肪酸穿过线粒体内膜需要载体肉毒碱转运,脂肪酸在血中转运需要与白蛋白结合生成脂酸白蛋白,而酮体通过线粒体内膜以及在血中转运并不需要载体。
(2)易利用:脂肪酸活化后进入β-氧化,每经4步反应才能生成一分子乙酰CoA,而乙酰乙酸活化后只需一步反应就可以生成两分子乙酰CoA,β-羟丁酸的利用只比乙酰乙酸多一步氧化反应。因此,可以把酮体看作是脂肪酸在肝脏加工生成的半成品。
(3)节省葡萄糖供脑和红细胞利用:肝外组织利用酮体会生成大量的乙酰CoA,大量乙酰CoA
抑制丙酮酸脱氢酶系活性,限制糖的利用。同时乙酰CoA还能激活丙酮酸羧化酶,促进糖异生。肝外组织利用酮体氧化供能,就减少了对葡萄糖的需求,以保证脑组织、红细胞对葡萄糖的需要。脑组织不能利用长链脂肪酸,但在饥饿时可利用酮体供能,饥饿5?周时酮体供能可多达70%.
(4)肌肉组织利用酮体,可以抑制肌肉蛋白质的分解,防止蛋白质过多消耗,其作用机理尚不清楚。
(5)酮体生成增多常见于饥饿、妊娠中毒症、糖尿病等情况下。低糖高脂饮食也可使酮体生成增多。