概述
P.马赫什
瓦里1950年根据
大孢子分裂次数、是否存在
核融合、成熟胚囊中细胞数目、排列和
染色体倍性等特征,把胚囊划分为10个类型:
蓼型、
月见草型、葱型、椒草型、贝母型、小白花丹型、德鲁撒型、五福花型、皮耐亚型、白花丹型,此外,根据核的数目和细胞排列的变异等,有人把胚囊分成11、13或16个类型。
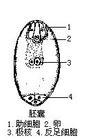
被子植物胚囊最普遍的形态是8核、7细胞的结构。其中卵是胚囊的重要组成部分。它与两个助细胞构成
卵器。
卵细胞在朝珠孔方向的壁较厚,朝
合点端的壁逐渐变薄,或者合点端没有壁。
中央细胞是胚囊中的最大细胞。其细胞壁从胚囊的珠孔端到
合点端,变化很大。中央细胞与卵、助细胞和反足细
 胚囊
胚囊
胞之间通过
胞间连丝联系。中央细胞核一般2个,称极核,位于细胞中央或靠近侧面的细胞质中。
反足细胞,一般在受精前后退化。有时会出现体积扩大和数目增多的现象。禾本科植物受精后反足细胞可发生一系列有丝分裂,形成几十个甚至多达几百个反足细胞。
大多数被子植物胚囊的整个表面具有吸收机能。可以消化邻接的
珠心细胞以致
珠被内层细胞。
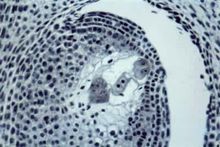
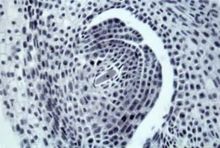
包藏于被子植物珠心中的一种高度特化的雌配子体,为
大孢子有丝分裂的产物(见彩图)。一般为 8核、7细胞的结构,包括一个
卵细胞、两个
助细胞、两个中央细胞核(
极核)和3个
反足细胞(图1)。受精后、受精卵在胚囊内发育成胚,受精的极核发育成
胚乳。
奥地利植物胚胎学家K.施拉夫1931年最先划分出不同的胚囊
 胚囊
胚囊
类型,继后1950年
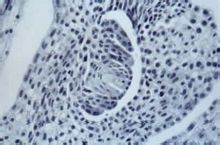
印度植物胚胎学家P.马赫什瓦里作了进一步的总结。胚囊发育包括
大孢子发生和雌配子体形成两个步骤。根据
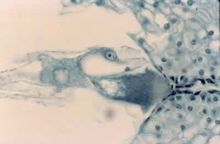
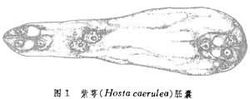
大孢子母细胞减数分裂后参加形成胚囊的大孢子核数目的不同,被子植物胚囊可分成单孢、双孢和四孢胚囊3种主要类型(图2)。单孢胚囊的特点是
大孢子母细胞经过减数分裂Ⅰ和Ⅱ产生4个线形排列的大孢子,其中3个退化,只有一个大孢子进一步发育成胚囊。双孢胚囊的特点是
大孢子母细胞进入减数分裂Ⅱ,另一个
二分体细胞退化。有功能的
二分体细胞在进行减数分裂Ⅱ时,不形成
细胞壁,这样产生的两个大孢子核都参与胚囊形成。四胞胚囊的特点是减数分裂Ⅰ与Ⅱ都不产生壁,结果形成4个
大孢子核,并全部参与
胚囊形成。由于
胚囊发育的细节有很大差异,因此上述3种主要类型的胚囊又可再分为若干不同类型。
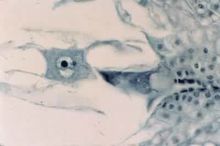
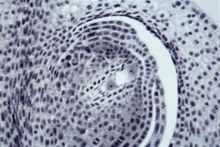
蓼型
由合点端有功能的一个大孢子经过 3次连续有丝分裂形成,
 胚囊
胚囊
最初8个核分为两群,每群4个核,一群在
胚囊的
珠孔端,另一群在合点端。然后,珠孔端那群产生构成
卵器的一个
卵细胞和两个助细胞,以及一个上极核;
合点端那群形成3个反足细胞和一个下极核。上、下极核都属于中央细胞。所以成熟胚囊为8核、7细胞的结构。被子植物大约有70%以上的科其胚囊属于这种发育类型。
月见草型
J.M.格尔茨1908年首次在月见草属中发现。它是由珠孔端的大孢子经过两次
有丝分裂后产生的,所以成熟的胚囊为4个
细胞结构。
葱型
双孢胚囊,是E.A.施特拉斯布格1879年最先在葱中发现的。它的发育特点是,两个大孢子核经过两次有丝分裂,形成与蓼型相似的8个胚囊。
四孢胚囊发育比较复杂,根据减数分裂后4个核的不同排列,可把它们分成3个组:第1组──贝母型,小白花丹型和德鲁撒型,4个大孢子呈1+3排列: 珠孔端1个,合点端3个。第2组──
五福花型,4个大孢子核呈2+2排列:
合点端和珠孔端各有2个大孢子核。第3组──皮耐亚型,
椒草型和
白花丹型,4个大孢子呈1+1+1+1排列,即胚囊的两核和两侧各有一个大孢子核。
贝母型
V.班巴基奥尼1928年在研究贝母属和百合属胚囊之后,发现减数分裂形成的4个大孢子核不直接进入8核阶段,而是代之以“次生四核阶段”。即4个大孢子核先形成1+3排列,
合点端3个核彼此紧密靠拢。当珠孔端那个核正常分裂时,
合点端3个纺缍体融合成一共同
纺锤体,形成一个3倍
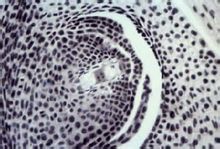 胚囊
胚囊
体核,然后再一分为二。结果,珠孔端两个
单倍体核,而
合点端两个为三倍体核。它们各自再分裂一次,才形成8核胚囊。但是,贝母型8核胚囊的核
倍性与正常蓼型胚囊不同,前者的下极核与反足细胞为三倍体,后者为
单倍体。
小白花丹型
是 F.法格林德和 J.W.博伊斯分别在1938和1939年研究小白花丹时发现的。它与
贝母型的区别在于,胚囊经“次生二核”后,终止在“次生四核”阶段。所以成熟胚囊只有4个核。
德鲁撒型
A.霍坎松1923年在伞形科的
Drusa op-
positifolia中发现了这一类型。其大
孢子形成阶段与贝母型相似。 但胚囊基部的3个大孢子核不融合。这样,1+3排列的核经过2次
有丝分裂,形成4+12排列的16核胚囊。
五福花型
是B.约恩松1879~1880年首先在五福花上描述的。其4个大孢子核经过一次
有丝分裂,就产生类似蓼型的8核胚囊。
皮耐亚型
是E.L.斯蒂芬斯1909年在皮耐亚科的皮耐亚属、
Brachysiphon属和
Sarcocolla属看到的,其胚囊发育方式非常特殊。4个大孢子核经过2次
有丝分裂,产生16核胚囊。
椒草型和皮耐亚型一样为16核胚囊,但成熟胚囊的细胞排列不一样。
白花丹型
是A.W.豪普特1934年描述的兰茉莉胚囊类型,它与皮耐亚属的主要区别在于:4个大孢子核只经一次有丝分裂,结果形成1个特殊的8核胚囊。
白花丹型胚囊特点是不具助细胞。

 胚囊
在裸子植物于胚乳动期从近珠孔的细胞分化为颈卵器。因此胚囊是由胚乳和颈卵器构成的单倍的配子体。或包藏于被子植物胚珠中的一种高度特化的雌配子体,为大孢子有丝分裂的产物。一般为"七胞八核"(即7个细胞,8个细胞核)的结构,包括一个卵细胞、两个助细胞、一个中央大细胞(中央大细胞内含有两个极核)和3个反足细胞。受精后,受精卵在胚囊内发育成胚,受精的极核发育成胚乳。正常型是在减数分裂形成的四个细胞中,有三个退化,剩下的含有一个核的胚囊细胞,核分裂三次,成为八个核,三个集中到近珠孔端,形成卵器,三个集中到近合点端,形成反足细胞,二个核移至胚囊中央形成极核。但是还
胚囊
在裸子植物于胚乳动期从近珠孔的细胞分化为颈卵器。因此胚囊是由胚乳和颈卵器构成的单倍的配子体。或包藏于被子植物胚珠中的一种高度特化的雌配子体,为大孢子有丝分裂的产物。一般为"七胞八核"(即7个细胞,8个细胞核)的结构,包括一个卵细胞、两个助细胞、一个中央大细胞(中央大细胞内含有两个极核)和3个反足细胞。受精后,受精卵在胚囊内发育成胚,受精的极核发育成胚乳。正常型是在减数分裂形成的四个细胞中,有三个退化,剩下的含有一个核的胚囊细胞,核分裂三次,成为八个核,三个集中到近珠孔端,形成卵器,三个集中到近合点端,形成反足细胞,二个核移至胚囊中央形成极核。但是还
 胚囊
有更复杂的形式,例如贝母型(Fritillaria-type)胚囊细胞为四核,其中近合点的三核分裂时与纺锤丝平行愈合成为3n的二核(将此称为Bambaccioni现象),不久同时分裂成为3n的四核,其中三个成为反足细胞,剩余的一个与近卵器的核(n)合成4n的中央核等各种形式,受精后,卵细胞成为二倍的而参与胚的形成。助细胞一般寿命短,在花粉管进入胚囊前后即行消失。反足细胞寿命也短,但其中也有在数量、大小上均增加而显有原叶体迹象的[龙胆科(Gentianaceae)、箬竹属(Sasa)等的几种]。此外,也有从卵细胞以外的细胞发育成胚的(多胚形成)。
胚囊
有更复杂的形式,例如贝母型(Fritillaria-type)胚囊细胞为四核,其中近合点的三核分裂时与纺锤丝平行愈合成为3n的二核(将此称为Bambaccioni现象),不久同时分裂成为3n的四核,其中三个成为反足细胞,剩余的一个与近卵器的核(n)合成4n的中央核等各种形式,受精后,卵细胞成为二倍的而参与胚的形成。助细胞一般寿命短,在花粉管进入胚囊前后即行消失。反足细胞寿命也短,但其中也有在数量、大小上均增加而显有原叶体迹象的[龙胆科(Gentianaceae)、箬竹属(Sasa)等的几种]。此外,也有从卵细胞以外的细胞发育成胚的(多胚形成)。