细胞表面的受体通过与其相应配体作用后,可经不同种类的G蛋白偶联,分别发挥不同的生物学效应。与G蛋白偶联的多种
受体具有共同的结构功能特点:分子量40-50kDa左右,由350-500
氨基酸组组成,形成7个由疏水氨基酸组成的
α螺旋区段,反复7次穿越细胞膜的脂质双层。
肽链的N末端在胞膜外,C末端在细胞内。N末端上常有许多
糖基修饰。从功能上看,
受体的识别区域并不象一般想象的那样在胞膜的外部,实际上是由7个跨膜区段间通过特定
氨基酸残基之间的
相互作用形成复杂的空间构象。配体结合于识别区域之后,即导致整个
受体构象的变化。受体
肽链的C末端和连接第5和第6个跨膜区段的第三个胞内环是G蛋白
结合部位。目前研究发现,趋化因子受体家族(chemokine receptor family)以及一些神经递质受体都属于G蛋白偶联的7次跨
膜受体的
超家族。例如IL-8RA胞膜外N端Asp11、Llu275、Arg280以及可形成
二硫键的Cys30和Cys277在与配体结合中起重要作用;紧接第三个空膜区第二个胞浆环中DRY序列对于与G蛋白的结合是必要的。
(1)Gs:
细胞表面受体与Gs(stimulating adenylate cyclase g protein,Gs)
偶联激活腺苷酸环化酶,产生cAMP
第二信使,继而激活cAMP依赖的
蛋白激酶。
(2)Gi:
细胞表面受体同Gi(inhibitory adenylate cyclase g protein,Gi)
偶联则产生与Gs相反的生物学效应。
(3)Gt:可以激活cGMP磷酸二酯酶,同视觉有关。
(4)Go:可以产生百日咳杆菌毒素导致的一系列效应。
(6)小G蛋白:近年来研究发现小G蛋白,特别是一些
原癌基因表达产物有着广泛的调节功能。Ras蛋白主要参与
细胞增殖和信号转导;
Rho蛋白对
细胞骨架网络的构成发挥调节作用;
Rab蛋白则参与调控
细胞内膜交通(membrane traffic)。此外,Rho和Rab亚家庭可能分别参与淋巴细胞极化(polarization)和抗原的提呈。某些信号蛋白通过SH-3功能区将酪氨酸激酶途径同一些由小G蛋白所控制的途径连接起来,如Rho(与Ras有30%同源性)调节胞浆中微丝上肌动蛋白的聚合或解离,从而影响细胞形态。这一事实解释了某些含有SH-3的蛋白同细胞骨架某些成份相关联或调节它们的功能。
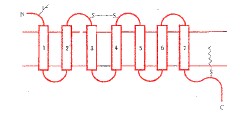 G蛋白偶联受体示意图(七个跨膜结构域)
G蛋白偶联受体示意图(七个跨膜结构域)

 G-蛋白概念的市场关注
G-蛋白概念的市场关注