淋巴系统
百科内容来自于:

概况
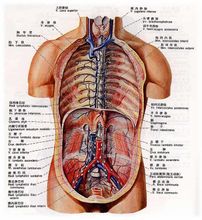
功能
淋巴管道

毛细淋巴管
毛细淋巴管lymphatiec capillary是淋巴管道的起始部,以膨大的盲端起始于组织间隙,收集多余的液体。其管壁由单层内皮细胞构成,内皮细胞间的间隙较大,无基膜和外周细胞,有纤维细丝牵拉,使毛细淋巴管处于扩张状态。因此毛细淋巴管壁的通透性较大,一些不易透过毛细血管的大分子物质,如蛋白质、细菌、异物、癌细胞等较易进入毛细淋巴管。毛细淋巴管分布广泛,除上皮、角膜、晶状体、牙釉质、软骨、脑和脊髓等处无毛细淋巴管外,遍及全身各处。
淋巴管
淋巴干
淋巴干 lymphatic trunk由淋巴管汇合而成。全身各部的浅、深淋巴管汇合成9条淋巴干 :收集头颈部淋巴的左、右颈干,收集上肢淋巴的左、右锁骨下干,收集胸部淋巴的左、右支气管纵隔干,收集下肢、盆部及腹部成对脏器淋巴的左、右腰干,收集腹部不成对脏器淋巴的肠干。
淋巴导管
1 胸导管thoraici duct 胸导管是全身最粗大的淋巴管道,长30-40cm。其下端起自乳糜池。乳糜池cisterna chyli 通常在第12胸椎下缘到第1腰椎体的前面,是由左、右腰干及肠干汇合而成的梭形膨大。胸导管起始后经主动脉裂孔入胸腔,沿脊椎右前方上行,至第5胸椎高度向左侧斜行,然后沿脊柱左前方上行,出胸廓上口至颈根部,呈弓形弯曲注入左静脉角。胸导管在注入静脉角之前还接纳左颈干、左左锁骨下干和左支气管纵隔干。胸导管收集双下肢、盆部、腹部、左半胸部、左上肢和左半头颈部的淋巴,即全身3/4部位的淋巴。
淋巴结
淋巴在向心流动中要通过一系列的淋巴结。淋巴结lymph nodes为圆形或椭圆开、大小不等的小体,一侧凸隆,另一侧凹陷,凹陷中央处为淋巴结门。与淋巴结凸侧相连的淋巴管称输入淋巴管,数目较多。出淋巴结门的淋巴管为输出淋巴管。淋巴结一般成群存在于较隐蔽的部位和胸、腹腔大血管附近。淋巴结的主要功能是滤过淋巴、产生淋巴细胞和参与免疫反应。了解局部淋巴结的位置、收集范围和引流去向,对临床诊断和治疗有一定意义。
全身各部的主要淋巴结
(二)上肢淋巴结:腋淋巴结
(三)胸部的淋巴结:支气管肺淋巴结,气管支气管淋巴结,气管旁淋巴结
(四)下肢的淋巴结:腹股沟浅淋巴结,腹股沟深淋巴结
(五)盆部的淋巴结:髂外淋巴结,髂内淋巴结,髂总淋巴结
(六)腹部的淋巴结:腰淋巴结,腹腔淋巴结,肠系膜上淋巴结,肠系膜下淋巴结
当病毒侵入人体发生感染时,淋巴结会肿大疼痛。像喉咙发炎时,会在下巴颏下摸到两个肿块,那就是淋巴结。炎症消失后淋巴肿块也会自然缩小。
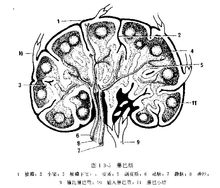
淋巴结的结构
淋巴结表面有被膜,被膜内面致密的部分称为皮质.中央部疏松的部分为髓质。
(1)被膜 淋巴结的被膜包绕在淋巴结皮质的表面,是由致密的结邹组织构成,在淋巴结的凸侧面有数条淋巴管穿入被膜、与被膜下淋巴窦相通。在淋巴结门有淋巴管、血管和神经出入。被膜中的结缔组织纤维束排列不规则,有些胶原纤维和弹件纤维束伸入淋巴结皮质与皮质中的纤维共同构成淋巴结皮质的小梁,形成淋巴结的网状支架。
(2)皮质 淋巴结的皮质位于被膜的下方,主要由间质性结缔组织和各种类型的细胞构成。皮质区有皮质淋巴窦、皮质区、副皮质区和淋巴小结区。
皮质淋巴窦包括被膜下窦、皮质小梁间淋巴窦和副皮质淋巴窦。这些淋巴窦互相通连并与髓质淋巴窦相通。各部位的窦壁均由淋巴内皮细胞构成,内皮细胞间连接松弛.有大小不等的间隙,细胞的结缔组织面有断续可见的稀薄的基膜,有些部位完全缺如。内皮细胞核呈不规则的长形,胞质中线粒体、糙面内质网较少,近管腔而有许多吞噬泡,偶尔可见脂类包含物。淋巴内皮细胞的胞质与胶原纤维和其它结缔组织密切接触。个别部位在电镜下可见到巨噬细胞通过内皮细胞间隙迁移。窦腔内含有大、小淋巴细胞、巨噬细胞、浆细胞;红细胞很罕见。巨噬细胞多靠近窦腔的边缘,其伪足常通过内皮细胞间隙伸至窦腔外的结缔组织中。
(3)髓质 髓质位于淋巴结的中央部.主要由髓索和髓质淋巴窦构成。髓索是由淋巴组织构成的索条状结构,相互连接成网。形状不规则.以网状纤维组织的支架,网间浆细胞和巨噬细胞占明显的优势。淋巴细胞和纤维母细胞较少、髓质区的浆细胞形状变化多端,糙面内质网丰富,带有粗糙的颗粒,呈环状,有些部位明显扩大成囊状膨大。细胞核附近有明显的高尔基氏器。髓索间的髓窦结构与皮质窦相似.并由皮质窦处延续而来、腔隙比皮质窦宽阔,形状不规则,由皮质窦流入髓窦的淋巴,最终汇入输出淋巴管、从淋巴结门穿出。髓质中有成团的巨噬细胞,浆细胞,可称为巨噬—浆细胞岛,其胞突相互交搭重叠、密切接触、也与其它淋巴组织细胞相联系,这些细胞也可与红细胞相接触。不同类型的细胞质突之间的接触联系,增强了细胞的吞噬免疫功能,尤其淋巴细胞、巨噬细胞可通过淋巴内皮细胞间隙迁移,有利于淋巴结的防御、免疫功能作用。
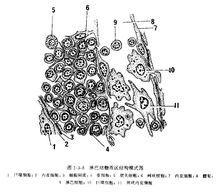
淋巴组织
淋巴组织是由网状细胞和网状纤维构成的结缔组织,又称为免疫组织。网状纤维形成细胞间微细的网状支架,主要有胶原纤维和弹件纤维构成。构成胶原纤维束的纤维轴直径约65nm,纤维束排列不规则,有些胶原纤维与弹性纤维相互交义。网状纤维间有大量的淋巴细胞,部分巨噬细胞,网状细胞。有散在的浆细胞和成纤维母细胞等。在不同部位的淋巴组织这些细胞的多少是有区别的,在炎性与非炎性区,这些细胞的比例有很显著性差别。如在炎症发作区域,淋巴细胞和巨噬细胞大量增加,巨噬细胞有活跃的伪足、吞噬活性增加。
淋巴组织在人体分布广泛.如呼吸道、消化道及尿生殖器的粘膜等处。它易与淋巴结.扁桃体.胸腺,脾等淋巴器官的构成。特别在空腔性器官的管腔大小和方向位置突然发生改变的部位,呼吸道的鼻咽部、咽峡、食管下端、胃喷门部、幽门部、十二指肠、回肠、阑尾、乙状结肠;输卵管的子宫部,静脉丛等部位都是淋巴组织较集中的地方。淋巴组织可分为弥散性淋巴组织和淋巴小结。
弥散淋巴组织
弥散性淋巴组织分布较分散,组织中的淋巴细胞、巨噬细胞和网状细胞也较稀疏,与相邻的组织没有明显的界限, 主要分布在消化道,呼吸道和尿生殖器官的粘膜下。部分出现在胸膜、腹膜下和淋巴管,小静脉的内皮下层。
淋巴小结
弥散的淋巴组织与淋巴小结严格说来没有明显的界限.随着机体的生理病理过程而有所改变。如弥散的淋巴组织,在有炎症情况下,淋巴细胞、巨噬细胞大量增加,有时可形成淋巴小结;有时淋巴小结中的淋巴细胞可以分散,稀疏配布.而变成弥散的淋巴组织,甚或变成为一般的网状结缔组织。
淋巴小结是淋巴细胞密集、形成具有一定形态和结构相似的淋巴组织团块,散在于淋巴组织之中,多呈圆形或卵圆形,直径在0.2-2mm。在淋巴结、胸腺、脾等淋巴器官中分布有许多淋巴小结。在消化道、呼吸道的粘膜下等处也分布有淋巴小结。单个分布的小结称为淋巴弧结,数个或数十个淋巴小结聚集成团构成淋巴集结。淋巴小结与淋巴结的区别在于淋巴小结没有输入淋巴管和淋巴窦。淋巴结产生的淋巴细胞一部分经上皮细胞间隙进入组织间隙。淋巴小结主要由B淋巴细胞组成,网状纤维组织构成了淋巴小结的微细支架,淋巴小结周围弥散的淋巴组织的网状纤维支架个主要有T淋巴细胞填允。但有时也有相当数量的浆细胞和巨噬细胞等。
淋巴系统的发生发展
淋巴系统的发生经历了漫长的衍化过程。原始动物到高等动物,脉管的通透性越来越低,在脊椎动物内皮细胞及其连接构成了通透屏障。而在无脊椎动物,这一屏障是由周皮细胞构成。在有关的比较解剖学研究中,常产生三种错误认识:(1)认为新一代原始动物是从他们相邻的前一类动物中分化而来的,忽视了它们在某个时期出现的飞跃突变发展的可能性。(2)忽视了动物也可以退化的事实。(3)认为相同的只会发生一次,忽视了集中进化。
人们研究了水蛭纲,发现这类寄生虫的脉管系统实际上是体腔发育衍化来的。其脉管有连续密闭的内皮细胞层,内皮细胞层外周并有不连续的周皮细胞,内皮细胞有许多窗孔。只有脊椎动物的脉管才具有这些结构特征。从脉管进化来看,水蛭似乎是所有无脊推动物中最高等的动物。并且认为水蛭的体腔脉管系统已与脊椎动物的淋巴系统相接近,甚于有报告发现水蛭有“淋巴心”的存在。
软体动物门的头足纲,通过对鱿鱼和章鱼的大量研究得出了基本相同的结论,其脉管壁通常具有一层完整的基膜和一层完整的内皮细胞,它们之间具有无脊椎动物类型的粘连小带连接,其间隙在20nn,间隙中充满一些可能作为通透屏障的电子密度较高的物质。但头足纲的脉管有多种类型,有的内皮细胞不连续,其间隙大小悬殊较大,细胞有许多突起,并含有一些囊泡、原纤维,且常有微管。某些部位如主动脉,有多层周皮细胞,肌层可见
内环层,外纵层的肌细胞,有收缩功能的血管,其周皮细胞内侧常有丝状纤维样结构。Andoyozhia(1953)认为头足纲已具有真正的淋巴系统,他将带染料的海水注射到套膜血管、发现有些脉管并未灌注显色,他认为这些未着色的脉管是真正的淋巴管。smith(1963)通过皮下注射和血管灌注TryPan蓝,未见有另一套脉管系统的存在。他认为前者的实验结果可能是由于海水引起部分脉管收缩从而未被灌注所致。他发现有些染料到达了肾囊.表明这
些染料颗粒是通过血管运输进入的。
对于腹足纲的脉管研究甚少,Amoroso等(1964)只发现其脉管有不连续的内皮细胞和连续的周细胞层。但没有发现明显的基膜。节肢动物的血循环系统大部分是开放的,心脏、大血管与整个体腔相通,所以脉管相对不发达。大量研究表明其管壁具有—层外膜,中间肌层和较厚的基膜。管腔内面是否存在内皮细胞,研究者各执己见,没有定论。一般认为存在不连续的内皮细胞.但其形态类似成纤维细胞,细胞质有许多突起伸入基膜。棘皮动物门有发育良好的水脉管系统,但它们的血管系统都发育很差,其管壁结构,细胞形态、基膜均与节肢动物门类似。对脊索动物门的脉管系研究一直被忽视,个别学者观察在大的血管的肌层内侧面有内皮细胞,但在小血管末发现有内皮细胞,仅有周细胞,脊索动物门更接近脊椎动物,其脉管系统基膜内面有内皮细胞,基膜外侧面有结缔组织。内皮细胞之间有裂隙,越到外周脉管,裂隙越多,越宽大,直到内皮细胞排列稀疏,最小的脉管仅由基膜构成,甚至某些部位没有基膜。这样血管直接与组织液接触。如果给动物喂以脂类,可以见到脂蛋白通过肠上皮,穿过基膜,通过内皮细胞间隙而进入血液。如果将墨汁注入体壁组织,碳粒很容易进入血管而被运送到全身。但脊索动物门有无真正的淋巴管,尚有争议。
到无颌类,血管系统已有相当大的进化.心脏和大血管壁的基膜外有相当厚的肌层。但足否有淋巴系统,仍有很大的争议。
到硬骨鱼类及更高级的动物的微血管与哺乳类基本相似,不再资述。内皮细胞的连接已十分紧密,血管系统再不能取代淋巴管的功能,随之建立了一个独立的淋巴系统。
两栖纲的淋巴系统,其最大特点是无尾类出现了巨大的淋巴囊,主要在皮下深都.整个躯体表层充满了淋巴液。而幼年无尾类及有尾类都无发达的淋巴囊。
随着动物的进化,脉管系统从体表到内部,变得越来越封闭。在无脊椎动物.周细胞起着密闭作用,在脊椎动物则出现了内皮细胞。在所有动物种系发生中,原始的外周循环系统与组织间隙相通联,并起着非常重要的作用。在较原始的动物,它们的开放性的血管相当于高等动物淋巴系的功能,同时辅以组织蛋白水解。在原始的脊椎动物,虽然具备淋巴系统的泵装置,但必须靠邻近肌肉的收缩才能推动淋巴循环。高级的脊椎动物有了淋巴心,而后整个收集淋巴管具有自身的收缩功能,再有淋巴瓣膜的配合,推动淋巴定向运行并注入血循环。同时也出现了具有滤过和免疫功能的淋巴结。
淋巴系统的研究方法
1、直接注射法
直接注射方法是将带色的注射剂,用注射针直接插入较粗大的淋巴管或淋巴结中.注入带色染料使淋巴管、淋巴结显色。最初应用水银淋巴管注射,仅适用于尸体淋巴管的解剖观察,现已很少应用,逐渐发展到应用油性和水溶性注射刑。
2、间接注射法
间接注射又称组织内注射,是将带色的注射剂注入组织或器官的组织间隙内,因为毛细淋巴管的内皮细胞间存在着开放间隙,借助于注射压力和注射液的扩散作用,带色的注射剂进入毛细淋巴管,使毛细淋巴管、集合淋巴管及淋巴结显示清楚。必要时,为了观察远距离的淋巴管和淋巴结,有时进行补充注射,使带色注射剂更好地充盈远处的淋巴管和淋巴结。
3、活体淋巴管显示方法
$firstVoiceSent
- 来自原声例句